
Porque no flexibilizar un músculo en contractura
Lic. Leandro Wachtmeister
Lic. Gabriel Pidello Rossi
RESUMEN
Considerando la importancia de una acción preventivo / terapéutica interdisciplinaria de las consecuencias funcionales sobre las estructuras musculares.
Objetivo: Estudiar la “Incidencia de la inhibición miotática de la neurona motora Beta y su implicancia en la Contractura Muscular”.
Método: Evaluación ecográfica de la condición muscular previa y post inhibición miotática en 8 deportistas profesionales entre los 18 a 30 años (media de 24 años).
Resultados: Nuestro interés en el estudio de la implicancia del sistema miotático Beta en el cuidado / protección del huso neuromuscular en la postura tónica del ser humano, nos llevó a profundizar el análisis sobre el momento del estiramiento muscular en el cual dicha neurona motora entra en función protectora. Nuestro objetivo es demostrar la incidencia de la inhibición miotática de la neurona motora Beta y su implicancia en la contractura muscular.
Para este estudio, se recurrió a las ecografías como forma de registro de los estados de la estructura muscular pre y post intervención sobre la contractura (estado de reposo y post inhibición del músculo) en 8 deportistas profesionales.
Las observaciones realizadas a partir de las bases neurofisiológicas y anatómicas del sistema miotático muestran que, si llevamos al sistema articular, mediante el sistema ligamentario y muscular, al máximo de estiramiento de la estructura muscular contracturada, retroalimentamos a la contractura ya engendrada y establecemos al mismo tiempo cambios en el Sistema Tónico Postural, generando cambios morfológicos en articulaciones. Esto ocurre cuando el huso neuromuscular – estirado más allá de sus límites fisiológicos – no se relaja dado que las partes polares continúan siendo estimuladas por la vía nociceptiva instalada. Esto continuara en estiramiento permanente de las partes centrales sensitivas que, por las fibras Ia, estimula permanentemente la motoneurona alfa, manteniendo así a las fibras musculares en contracción permanente, que se exacerba llevando al músculo al máximo de estiramiento.
De los resultados, observamos una disminución del diámetro del músculo con un nivel de significación del 0,0469 en relación al músculo antes y después de la inhibición muscular. Las interpretaciones de los datos nos llevan a considerar que tiene una alta (95,31%) implicancia la neurona motora Beta en la contractura muscular.
Estas observaciones son de interés para la orientación de futuros trabajos de investigación que profundicen en los indicios aportados por nuestro estudio particularmente con relación al análisis de los efectos negativos en el trabajo de estiramientos de la estructura muscular en músculos en contractura.
INTRODUCCIÓN
Numerosas publicaciones describen los beneficios de los trabajos de estiramiento sin tener en cuenta el estado de la estructura muscular.
Nuestro interés en el estudio de la implicancia del sistema miotático Beta en el cuidado / protección del huso neuromuscular en la postura tónica del ser humano, nos llevó a profundizar el análisis sobre el momento del estiramiento muscular en el cual dicha neurona motora entra en función protectora.
Nuestro objetivo es demostrar la incidencia de la inhibición miotática de la neurona motora Beta y su implicancia en la contractura muscular.
PACIENTES, MATERIAL Y MÉTODOS
La población objeto de estudio está constituida por 8 deportistas profesionales comprendidos entre los 18 a 30 años (media de 24 años) que aceptaron voluntariamente participar en el estudio.
Fig. 1 Ecografía del músculo a evaluar. Procedimiento habitual para una ecografía muscular
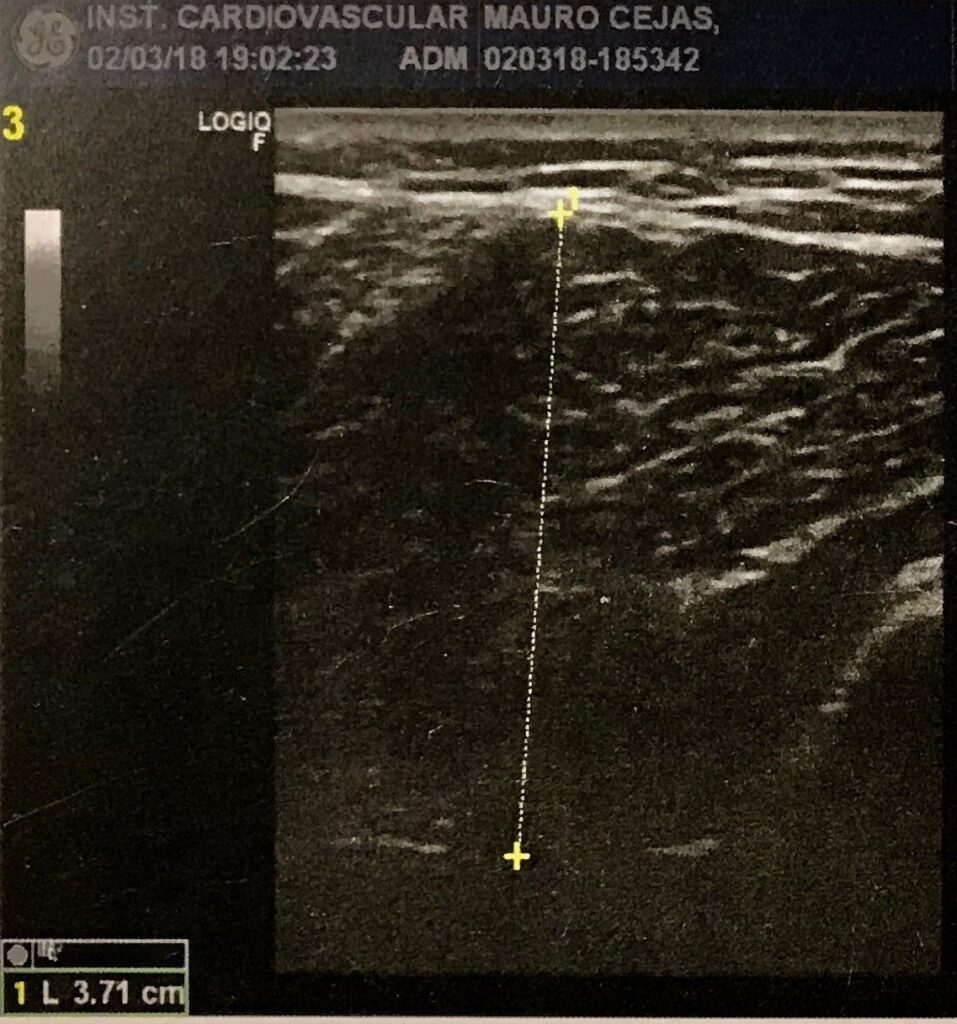
Fig. 2 Ecografía del músculo después del proceso de inhibición
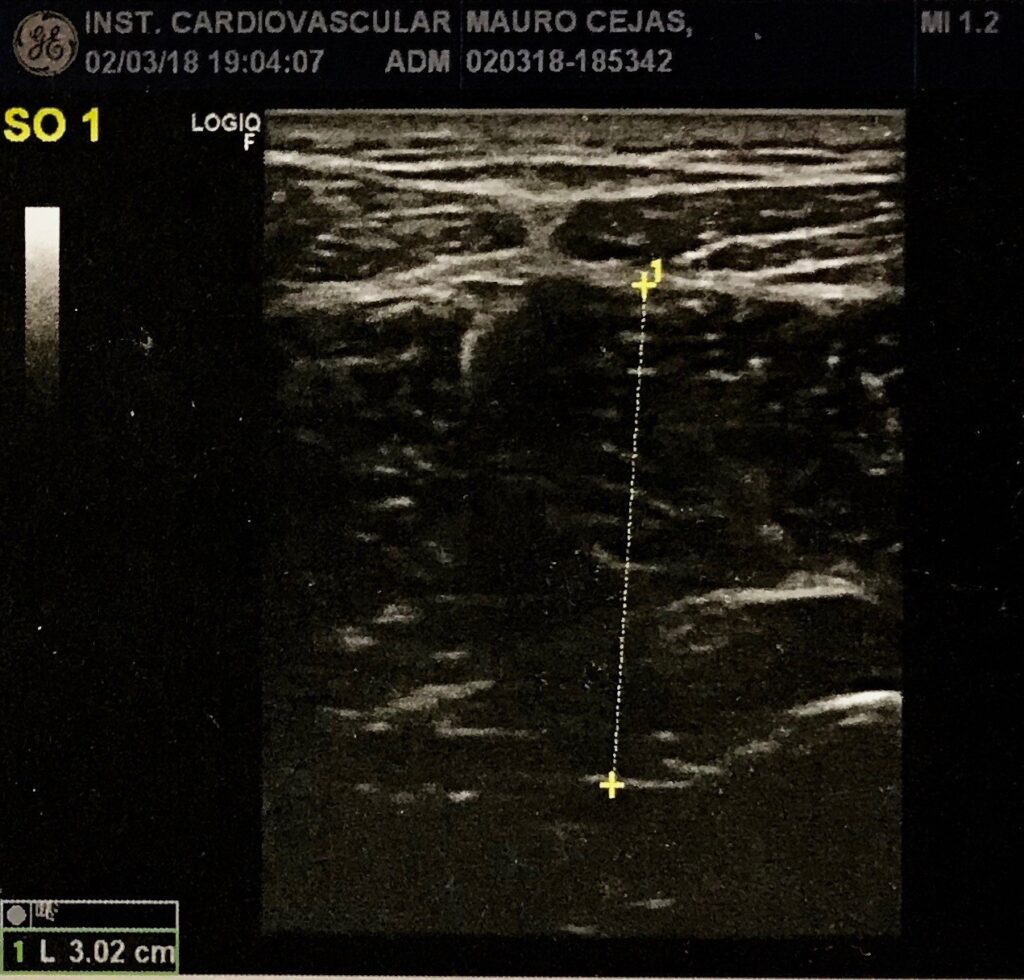
Para nuestro estudio, recurrimos a las ecografías como forma de registro de los estados de la estructura muscular pre y post intervención sobre la contractura.
DISCUSIÓN
En primer lugar, debemos señalar que, a pesar del creciente interés por determinar el origen de la contractura muscular dada su importancia en el ámbito terapéutico, actualmente se carece de estudios realizados sobre una sólida base metodológica.
Distintas teorías explican la estrecha relación entre las neuronas motoras alfa, gamma y no se cuenta con evidencias científicas sobre el comportamiento de la Neurona Motora Beta. Analizando los fundamentos Neurofisiológicos, nos basamos en la neurofisiología muscular que sostiene que en el músculo y las articulaciones existen receptores sensoriales de relación funcional y que el músculo como las articulaciones contienen receptores que envían al SNC información sobre: Tensión, Longitud, Presión y nocicepción. Dos de estos receptores musculares tienen especial significado para la actividad contráctil: el Huso neuromuscular y el órgano tendinoso de Golgi (GTO). Los husos musculares lo integran fibras musculares especiales “intrafusales”, terminaciones nerviosas sensoriales y motoras que regulan la excitabilidad de las fibras nerviosas sensoriales (aferentes), a través de las deformaciones mecánicas de las superficies receptora de esas fibras. Las fibras intrafusales reciben inervación motora de un grupo de motoneuronas pequeñas llamadas motoneuronas Gamma que forman partes del pool de motoneuronas del asta anterior que inerva a dicho músculo. Las fibras “extrafusales” son las que dan fuerza contráctil al músculo y están inervadas por un grupo de motoneuronas grandes (motoneuronas Alfa). Un tercer tipo de inervación muscular, llamadas Beta o esqueleto – fusimotoras inervan tanto fibras musculares intrafusales como extrafusales. Las motoneuronas Beta difieren de las gammas, además de su tamaño, en que las Beta participan en el estiramiento (miotático). Esta diferencia es importante funcionalmente, ya que en respuesta al estiramiento muscular las motoneuronas Beta producen la coactivación de las fibras extrafusales e intrafusales y así preservan la actividad de las fibras aferentes tipo Ia . Existen además 2 tipos de terminaciones aferentes (sensoriales) en el huso muscular: Primarias (fibras nerviosas tipo I que inervan cada fibra intrafusal, formando un espiral alrededor de la fibra; y Secundarias (fibras nerviosas tipo II) que inervan de manera preponderante las fibras extrafusales. “Ambos tipos sensoriales descargan información cuando se estira el músculo”. Los órganos tendinosos de Golgi (GTO) son estructuras presentes en el tendón. No tienen, a diferencia de los husos musculares, inervación motora, pero presentan una rica inervación sensorial por fibras de tipo Ib, que descargan tanto cuando el músculo se contrae de forma concéntrica como excéntrica.
Al registrar la actividad eléctrica de las fibras aferentes del huso muscular y del órgano tendinoso, el análisis de las imágenes de los diferentes estados que atraviesa (pasa) la condición de la estructura muscular muestra que:
- En estado de Reposo: solo descarga el huso muscular.
- Ante el estiramiento muscular – momento de la resistencia es mayor al momento de la fuerza – (fuerza excéntrica): ambos receptores descargan, el huso muscular y el órgano tendinoso.
- En la contracción muscular, (fuerza isométrica) solo el órgano tendinoso descarga.
- Para poder aplicar este concepto en la terapéutica, es necesario establecer si el músculo se encuentra en contractura (desarrollado más adelante de este artículo) o se encuentra en acortamiento miofascial.
- Durante la contracción muscular, el huso aumenta su longitud relativa en relación con la fibra muscular extrafusal contraída y, por consiguiente, se pliega y deja de descargar.
Sabemos que la activación simultánea de motoneuronas alfa y gamma (coinervadas por Beta) tienen como finalidad que el huso muscular siga cumpliendo su función de receptor, proveyendo información estática y dinámica sobre la longitud muscular. Es con relación a esta función que planteamos la necesidad de conocer la importancia de la activación de la motoneurona Beta que en forma simultánea inerva a las fibras intrafusal y extrafusal.
Para comprobar esta hipótesis tuvimos que establecer primeramente si el sistema muscular se encuentra en contractura o se encuentra en acortamiento fascial producto de la disfunción del sistema fascial.
A partir de lo desarrollado sobre las bases neurofisiológicas y anatómicas del sistema miotático, llegamos a la discusión, de que, si llevamos al sistema articular, mediante el sistema ligamentario y muscular, al máximo de estiramiento de la estructura muscular contracturada, retroalimentamos a la contractura ya engendrada y establecemos al mismo tiempo cambios en el Sistema Tónico Postural, generando cambios morfológicos en articulaciones. Esto ocurre cuando el huso neuromuscular – estirado más allá de sus límites fisiológicos – no se relaja dado que las partes polares continúan siendo estimuladas por la vía nociceptiva instalada. Esto continuara en estiramiento permanente de las partes centrales sensitivas que, por las fibras Ia, estimula permanentemente la motoneurona alfa, manteniendo así a las fibras musculares en contracción permanente, que se exacerba llevando al músculo al máximo de estiramiento.
Posteriormente tratamos la contractura, la cual se inhibió el reflejo miotático beta en el sistema nervioso, más precisamente en la formación bulboreticular, y de esa manera desactivar el bucle patológico – anormal establecido, luego se realizó una nueva ecografía.
RESULTADOS
Se aplica el Test de aleatorización de Fisher para dos muestras relacionadas a los resultados de las evaluaciones ecográficas. El análisis de las variaciones observadas es el que sigue:
¡Se consideran 2! ×n = 2 × 8 = 16 individuos que están relacionados y designa con:
X! 1i: Las observaciones del grupo sometido al Tratamiento 1 (Reposo).
X! 2i: Las observaciones del grupo sometido al Tratamiento 2 (Inhibición).
Y las diferencias de los n! pares: d! i = X1i −X2i
Las observaciones deben ser medidas en al menos una escala de intervalo. El método no requiere supuesto alguno sobre las distribuciones de las variables aleatorias bajo estudio.
Se desea ensayar si después de aplicar el Tratamiento 2 se obtuvo una disminución significativa del diámetro del músculo, es decir: μ! 1 > μ2
Por lo tanto las hipótesis a ensayar son:
H! 0)μ1 = μ2
H! 1)μ1 > μ2
| Par | Tratamiento 1 | Tratamiento 2 | Diferencia !(di) |
| 1 | 3.20 | 3.02 | 0.18 |
| 2 | 3.40 | 3.41 | -0.01 |
| 3 | 3.17 | 3.00 | 0.17 |
| 4 | 0.45 | 0.40 | 0.05 |
| 5 | 0.33 | 0.21 | 0.12 |
| 6 | 3.43 | 3.39 | 0.04 |
| 7 | 3.37 | 3.14 | 0.23 |
| 8 | 3.20 | 2.97 | 0.23 |
Bajo la hipótesis nula cada par tiene la misma probabilidad de que d! i > 0 ó d! i < 0
Entonces, bajo estas condiciones, se podrían presentar otros resultados con algunas d! i de signo cambiado, y con igual probabilidad de ocurrencia, como consecuencia de distintas asignaciones de tratamientos.
El número total de formas distintas de asignar aleatoriamente los dos tratamientos en n! pares, está dado por Arreglos con Repetición de 2! elementos tomados de a n! .
A! ´2,n = 2n = 28 = 256
La probabilidad de cada asignación es 1! /256 = 0.0039
La estadística a emplear para ensayar la hipótesis nula, es la suma de las diferencias.
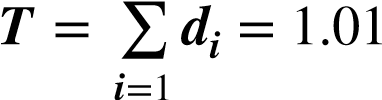
Se rechaza la hipótesis nula cuando T! toma valores significativamente extremos. Si para cada asignación aleatoria de los tratamientos se calcula el valor de T! , se obtiene la Distribución completa de H! 0 . Para evitar este trabajo, se calcula solamente los valores más extremos de T! , y en una cantidad equivalente a la proporción α! del total de valores posibles, determinando así la Región de Rechazo de tamaño α! .
K! = α*2n = 0.05*256 = 12.8
Se considera a k! = 12, por lo tanto resulta α! = 0.0469. Entonces se calculan los 12 valores de T! más extremos en sentido positivo.
| Asignación 1 | Asignación 2 | Asignación 3 | Asignación 4 | Asignación 5 | Asignación 6 |
| 0.18 | 0.18 | 0.18 | 0.18 | 0.18 | 0.18 |
| 0.01 | -0.01 | 0.01 | 0.01 | 0.01 | 0.01 |
| 0.17 | 0.17 | 0.17 | 0.17 | 0.17 | -0.17 |
| 0.05 | 0.05 | 0.05 | -0.05 | 0.05 | 0.05 |
| 0.12 | 0.12 | 0.12 | 0.12 | -0.12 | 0.12 |
| 0.04 | 0.04 | -0.04 | 0.04 | 0.04 | 0.04 |
| 0.23 | 0.23 | 0.23 | 0.23 | 0.23 | 0.23 |
| 0.23 | 0.23 | 0.23 | 0.23 | 0.23 | 0.23 |
| T= 1.03 | T= 1.01 | T= 0.95 | T= 0.93 | T= 0.79 | T= 0.69 |
| Asignación 7 | Asignación 8 | Asignación 9 | Asignación 10 | Asignación 11 | Asignación 12 |
| -0.18 | 0.18 | 0.18 | 0.18 | 0.18 | 0.18 |
| 0.01 | 0.01 | 0.01 | -0.01 | 0.01 | 0.01 |
| 0.17 | 0.17 | 0.17 | 0.17 | 0.17 | 0.17 |
| 0.05 | 0.05 | 0.05 | 0.05 | -0.05 | 0.05 |
| 0.12 | 0.12 | 0.12 | 0.12 | 0.12 | -0.12 |
| 0.04 | 0.04 | 0.04 | 0.04 | 0.04 | 0.04 |
| 0.23 | -0.23 | 0.23 | 0.23 | 0.23 | 0.23 |
| 0.23 | 0.23 | -0.23 | -0.23 | -0.23 | -0.23 |
| T= 0.67 | T= 0.57 | T= 0.57 | T= 0.55 | T= 0.47 | T= 0.33 |
Como T! = 1.01 > 0.33, por lo tanto se rechaza la hipótesis nula y se puede concluir con un nivel de significación del 0.0469! que se obtuvo una disminución del diámetro del músculo.
CONCLUSIÓN
El problema que se planteó en este estudio fue analizar la “Incidencia de la inhibición miotática en la neurona motora Beta y su implicancia en la Contractura Muscular”
Para tratar de dar respuesta a este estudio, se evaluaron 8 deportistas profesionales comprendidos entre los 18 a 30 años (media de 24 años), a los que se le realizó una ecografía en reposo, y luego otra ecografía después de la inhibición del músculo.
De los resultados, observamos una disminución del diámetro del músculo con un nivel de significación del 0,0469 en relación al músculo antes y después de la inhibición muscular. Las interpretaciones de los datos nos llevan a considerar que tiene una alta (95,31%) implicancia la neurona motora Beta en la contractura muscular.
Estas observaciones son de interés para la orientación de futuros trabajos de investigación que profundicen en los indicios aportados por nuestro estudio particularmente con relación al análisis de los efectos negativos en el trabajo de estiramientos de la estructura muscular en músculos en contractura.
También te puede interesar

Organización Estructural del Movimiento
